

In PVLG
|
|
In PVLG |
|
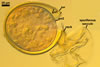
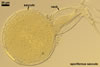
SPORES origin inside the neck of a sporiferous saccule at a 20-70(-100) µm distance from the saccule forming terminally from or intercalary inside mycorrhizal extraradical hyphae by swelling. Spores occur singly in the soil or in roots. Spores subhyaline when juvenile, pale yellow (4A3) to deep yellow (4A8) at maturity; globose to subglobose, ellipsoid to irregular; 85-140 x 95-210 µm.
 |
 |
In PVLG+Melzer's reagent |
|
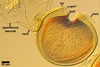
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germination walls.
Spore wall composed of three layers (swl1-3).
 |
 |
 |
 |
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's reagent |
|||||||
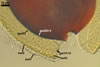
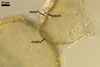
Layer 1, forming the spore surface, evanescent, (0.5-)0.8(-1.2) µm thick, continuous with the wall of the sporiferous saccule, usually present in spores with the sporiferous saccule attached, but sloughed when the saccule is detached or highly deteriorated.
Layer 2 laminate, pale yellow (4A3) to deep yellow (4A8), (2.9-)3.4(-4.4) µm thick, ornamented with pits; pits circular to subcircular, (1.0-)1.6(-2.8) µm diam, ellipsoid to irregular, 1.5-1.8 x 2.7-3.4 µm when seen in a plane view, (1.2-)1.6(-2.0) µm deep when observed in a cross sectional view, separated by ridges, (1.0-)1.8(-2.2) µm wide.
Layer 3 flexible to semiflexible, pale yellow (4A3) to deep yellow (4A8), (0.5-)0.7(-0.9) µm thick, always tightly adherent to the lower surface of layer 2 and, thereby, difficult to detect.
Germination wall 1 composed of two flexible to semiflexible, hyaline layers (gw1l1 and 2), usually not separating from one another; layer 1 usually slightly thinner, ca. 0.5-0.8 µm thick, and more flexible than layer 2, which is ca. 0.5-1.0 µm thick.
Germination wall 2 comprises three hyaline layers (gw2l1-3).
Layer 1 0.5-0.8 µm thick, covered with fine granular excrescences (beads), usually more or less dislocated in crushed spores.
Layer 2 plastic, 4-12 µm thick in lactic acid-based mountants.
Layer 3 very thin, <0.5 µm thick, rarely separating from layer 2 and, hence, exceptionally difficult to see.
In Melzer's reagent, only layer 2 of the germination wall 2 stains pale red (11A3) to madder red (11B8).
GERMINATION ORB. Not found.
GERMINATION. Unknown to date. Morton (2002) assumed K. kentinensis spores to germinate by a germ tube arising from a hyaline germination orb earlier formed on the second germination wall.
 |
 |
 |
 |
In PVLG |
|||
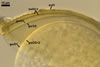
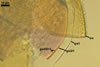
CICATRICES. Two circular scars are present. A scar proximal to the saccule is (20.0-)25.3(-32.0) µm diam, and that distal to the saccule is (5.0-)7.1(-10.5) µm diam when observed in a plane view. The cicatrices form after the closure of two opposite pores present in the structural layer (laminate spore wall layer 2) of a developing spore by sublayers (laminae) added to the lower surface of this layer and the spore wall layer 3. Both pores become closed after the full differentiation of the spore wall and before the beginning of the synthesis of the first germination wall (Sieverding and Oehl 2006).
SPORIFEROUS SACCULE consists of a saccule and a neck. Saccule pale yellow (4A3) in youth, hyaline at spore maturity; globose to subglobose; 100-150 µm diam; collapsed or detached in mature spores.
 |
 |
 |
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's reagent |
||||||
Wall of saccule composed of an evanescent, flexible, hyaline, 0.5-1.0 µm thick, outer layer (sswl1) and a semipermanent, semiflexible, hyaline to yellowish white (4A2), 0.5-1.2 µm thick, inner layer (sswl2).
Saccule neck hyaline to deep yellow (4A8); cylindrical or slightly gradually narrowing from the saccule to the spore; (20.0-)25.5(-37.5) µm wide at the saccule, (27.5-)31.8(-35.5) µm wide at the spore, then much narrower, (5.0-)7.5(-11.5) µm wide at the distal cicatrix and gradually tapering up to 2-3 µm wide. Wall of saccule neck consists of a hyaline layer (ssnwl1) continuous with the wall of the saccule and frequently of a pale yellow (4A3) to deep yellow (4A8) layer (ssnwl2) continuous with the laminate spore wall layer, extending up to 10-40(-100) µm into the lumen of the sporiferous saccule neck.
MYCORRHIZAE. According to Wu et al. (1995a, b), Morton (2002), and Sieverding and Oehl (2006), K. kentinensis formed mycorrhizae with arbuscules and vesicles staining intensively in trypan blue.
PHYLOGENETIC POSITION. So far, there is no literature report of the phylogenetic position of K. kentinensis.
DISTRIBUTION. According to Wu et al. (1995b), the holotype of K. kentinensis was grown in a pot culture with Paspalum notatum Flügge as the host plant (culture no. SM-22) at Taiwan Agricultural Research Institute, Wu-feng, Taichung. This fungus was originally collected from the rhizosphere of Eleusine indica (L.) Goertn. and Setaria viridis (L.) Baeuv. growing in bamboo vegetation (Phyllostachys pubescens Mazel) at Chi-tou, Taiwan. Additionally, spores of K. kentinensis have been found in Brazil (Carponi et al. 2003; Dos Anjos et al. 2005), Benin ( Western Africa; Sieverding and Oehl 2006), and Japan (Wu et al. 1995).
NOTES. The characterization of spores of K. kentinensis presented above was made based on the original description of this fungus (Wu et al. 1995b), the description emended by Sieverding and Oehl (2006), and results of examination of spores provided by Dr. F. Oehl, Institute of Botany, University of Basel, Switzerland; slides no. .......
Most properties of spores of K. kentinensis presented above agree with those of this fungus originally described by Wu et al. (1995b), and especially with those emended by Morton (2002) and Sieverding and Oehl (2006). The author of this website found only small differences in properties of the germination wall 1 and the wall of the sporiferous saccule.
According to Morton (2002), the germination wall 1 of K. kentinensis spores consists of two layers of a near equal thickness. In spores examined by me, the first layer of this wall always was thinner and more flexible than layer 2. Morton (2002) described the wall of the sporiferous saccule to consist of one layer, although the included microphotographs show two labeled layers of characters identical to those presented above.
Apart from K. kentinensis, the only other member of the genus Kuklospora is K. colombiana (Oehl and Sieverding 2006). Both fungi are easy to distinguish. While spores of K. kentinensis are ornamented with pits, those of K. colombiana are smooth (Blaszkowski, pers. observ.; Sieverding and Oehl 2006; Wu et al. 1995b). Additionally, spores of the former species may be markedly larger (up to 210 µm diam; Wu et al. 1995b) than those of the latter fungus (up to 135 µm diam; Schenck et al. 1984).
When devoid of sporiferous saccules, spores of K. kentinensis are almost indistinguishable from those of Acaulospora scrobiculata lacking sporiferous saccules. Spores of the two species are similarly coloured, overlap in the range of their diameter, and their structural laminate wall layer is ornamented with pits of a similar size and appearance (Blaszkowski, pers. observ., 2003; Morton 2002; Trappe 1977; Wu et al. 1995b). Moreover, the phenotypic and biochemical properties of their both germination walls are identical. The only character distinguishing spores of these fungi is the number of cicatrices: two in spores of E. kentinensis vs. one in those of A. scrobiculata).
REFERENCES
Carponi A. C., Franco A. A., Berbara R. L. L., Trufem S. B., Granha J. R. D., Monteiro A. B. 2003. Arbuscular mycorrhizal fungi occurrence in revegetated areas after bauxite mining at Porto Trometas, Pará State, Brazil. Pesq. Agropec. Bras. 38, 1409-1418.
Dos Anjos E. C. T., Cavalcante U. M. T., Dos Santos V. F., Maia L. C. 2005. Production of mycorrhized sweet fruit seedlings in disinfected and phosphorous fertilized soil. Pesq. Agropec. Bras. 40, 345-351.
Morton J. B. 2002. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University. http://www.invam.caf.wvu.edu/.
Schenck N. C., Spain J. L., Howeler R. H. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76, 685-699.
Sieverding E., Oehl F. 2006. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. Food Qual. 80, 69-81.
Trappe J. W. 1977. Three new Endogonaceae: Glomus constrictus, Sclerocystis clavispora, and Acaulospora scrobiculata. Mycotaxon 6, 359-366.
Wu C.-G., Liu Y.-S., Hung L. L. 1995a. Spore development of Entrophospora kentinensis in an aeroponic system. Mycologia 87, 582-587.
Wu C.-G., Liu Y.-S., Hwuang Y.-L., Wang Y.-P., Chao C.-C. 1995b. Glomales of Taiwan: V. Glomus chimnobambusae and Entrophospora kentinensis, spp. nov. Mycotaxon 53, 283-294.