


In PVLG
In PVLG+Melzer's
Spain & N.C. Schenck
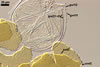
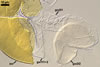
SPORES single in the soil; develop laterally on the neck of a sporiferous saccule; pale yellow (3A3) to orange (5B8); globose to subglobose; (98-)116(-140) µm diam; sometimes ovoid; 100-120 x 110-140 µm.
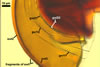
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germination walls.
Spore wall composed of three layers (swl1-3). |
|
|
|
|
|
In PVLG |
In PVLG+Melzer's |
|||
Layer 1 evanescent, hyaline, (0.8-)1.0(-1.3) µm thick, continuous with the wall of a sporiferous saccule, usually completely sloughed in mature spores.
Layer 2 laminate, pale yellow (3A3) to orange (5B8), (2.2-)2.8(-3.9) µm thick.
Layer 3 semiflexible, hyaline, (0.5-)0.7(-0.8) µm thick, rarely separable from layer 2.
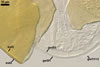
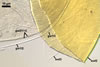
Germination wall 1 consists of two tightly adherent hyaline, semiflexible layers (gw1l1 and 2), each 0.4-0.6 µm thick; these layers sometimes separate from each other in crushed spores.
 |
 |
 |
 |
 |
 |
 |
In PVLG
|
||||||
Germination wall 2 composed of two adherent layers (gw2l1 and 2).
 |
 |
 |
 |
 |
 |
 |
In PVLG+Melzer's
reagent
|
||||||
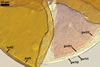
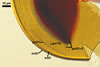
Layer 1 flexible, hyaline, (0.5-)0.9(-1.0) µm thick, covered with small, <0.5 µm diam, granules easily scattering in crushed spores.
Layer 2 plastic, hyaline, 10-12.5 µm thick in PVLG, (0.8-)1.1(-1.2) µm thick and beetroot purple (13D8) in Melzer’s reagent.
 |
 |
 |
In PVLG
|
||
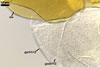
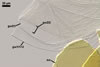
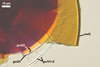
GERMINATION ORB hyaline to pale yellow (3A3); circular or somewhat elliptic; 60-62.5 x 65.0-70.0 µm when seen in a plane view; formed by a centrifugally rolled hypha, (7.1-)9.5(-12.0) µm wide, sometimes broadened up to 20.8 µm wide at the end. A circular germ tube initial, (2.1-)2.9(-3.4) µm diam is present at some distance from the hyphal end . The germination orb is positioned on the surface of germination wall 2.
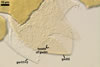
SPORIFEROUS SACCULE hyaline; globose to subglobose; 90-130 µm diam, with 1-layered wall, 0.8-1.2 µm thick; neck 50-80 µm long, tapering from 17.0-24.0 µm wide at the saccule to 15.0-20.0 µm wide at the spore attachment. Saccule usually collapses or is detached in mature spores.
 |
In PVLG |
CICATRIX. A slightly raised collar, 8.1-10.5 wide x 3.7-5.0 µm long, surrounding a hole 7.5-9.8 µm diam.
MYCORRHIZAE. In Poland, Acaulospora mellea has been associated in the field with roots of Ammophila arenaria (L.) Link, Corynephorus canescens (L.) P. Beauv., Crataegus monogyna Jacq., Elymus arenarius L., Festuca arundinacea Schreb., F. rubra L. s. s., Hieracium umbellatum L., Holcus mollis L., Hordeum vulgare L., Juncus balticus Willd., Juniperus communis L., Leontodon autumnalis L., Lupinus angustifolius L., Poa pratensis L., Rosa canina L., Triticum aestivum L., Verbascum phlomoides L., XTriticosecale Wittmack, and Vicia sativa L. (Blaszkowski 1993a, b, 1994; Blaszkowski et al. 2002).
In one-species pot cultures with Sorghum sudanense (Piper) Stapf, Ac. mellea formed mycorrhizae with arbuscules, vesicles, as well as intra- and extraradical hyphae staining intensively in 0.1% trypan blue (Blaszkowski et al. 1999).
DISTRIBUTION. In Poland, Ac. mellea has been found in ca. 40 soil samples. This fungus has occurred among roots of wild and cultivated plants growing in forest nurseries, uncultivated and cultivated soils (Blaszkowski 1993a), as well as in maritime (Blaszkowski 1993b, 1994) and inland (Blaszkowski et al. 2002) sand dunes.
Acaulospora mellea is probably a widely distributed arbuscular mycorrhizal fungus in the world. It has previously been recorded in cultivated and uncultivated soils of Florida, Massachusetts, North Carolina, Rhode Island, U. S. A. (Bever et al. 1996; Douds and Schenck 1990; Koske and Gemma 1997; Schenck et al. 1984; Sylvia 1986), Brazil (Grandi and Trufem 1991; Schenck et al. 1984), Mexico (Estrada-Torres et al. 1992), Colombia (Dodd et al. 1990; Saif 1987; Sieverding 1989; Sieverding and Toro 1988; Sieverding et al. 1986), Cameroon (Musoko et al. 1994), and China (Mei-ging and You-shan 1992).
NOTES. Spores of Acaulospora mellea may easily be confused with those of Ac. morrowiae Spain et N.C. Schenck due to the similarity in spore size and the structure of its spore wall and two germination flexible walls. The spores of Ac. mellea compared with those of Ac. morrowiae are usually somewhat larger [average 116 µm diam (pers. observ.), 120 µm diam (Morton 2000) vs. 60-100 µm diam (Morton 2000)] and darker-coloured [pale yellow to orange (pers. observ.), pale orange-brown to dark orange brown (Morton 2000) vs. subhyaline to pale yellow-brown (Morton 2000)].
Another arbuscular mycorrhizal fungus resembling Ac. mellea is Ac. gedanensis Blaszk. However, the latter species forms markedly smaller spores (55-88 µm in diam) of different properties of the layers of its both germination walls (Blaszkowski 1988, 1994). Compared with the two adherent semi-rigid layers of the germination wall 1 of Ac. mellea spores, the germination wall 1 of Ac. gedanensis spores is represented by a single, rigid layer, easily cracking in crushed spores. Additionally, in contrast to the beaded, flexible layer adherent to a highly plastic amorphous layer of the germination wall 2 of Ac. mellea spores, the germination wall 2 of Ac. gedanensis spores consists of two smooth, semirigid layers. None of these layers stains in Melzer’s reagent (vs. dark staining reaction in Ac. mellea).
The spore wall of Ac. gedanensis consists of an evanescent layer adherent to a yellow to pale brown laminate layer. According to Morton (2000), the laminate layer of probably all Acaulospora spp. is associated with a tightly adherent, thin, flexible layer. This frequently makes difficult or impossible to recognize the layer in many species of this genus, including Ac. gedanensis.
According to Morton (2000), Ac. mellea probably is synonymous with Ac. dilatata J.B. Morton.
REFERENCES
Bever J. D., Morton J. B., Antonovics J., Schultz P. A. 1996. Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. J. Ecol. 84, 71-82.
Blaszkowski J. 1988. Four new species of the Endogonaceae (Zygomycotina) from Poland. Karstenia 27: 37-42.
Blaszkowski J. 1993a. Comparative studies of the occurrence of arbuscular fungi and mycorrhizae (Glomales) in cultivated and uncultivated soils of Poland. Acta Mycol. 28, 93-140.
Blaszkowski J. 1993b. The occurrence of arbuscular fungi and mycorrhizae (Glomales) in plant communities of maritime dunes and shores of Poland. Bull. Pol. Ac. Sci. Biol. Sci. 41, 377-392.
Blaszkowski J. 1994. Arbuscular fungi and mycorrhizae (Glomales) of the Hel Peninsula, Poland. Mycorrhiza 5, 71-88.
Blaszkowski J., Tadych M., Madej T. 2002. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of the Bledowska Desert, Poland. Acta Soc. Bot. Pol. 71, 71-85.
Blaszkowski J., Tadych M., Madej T., Adamska I., Czerniawska B., Iwaniuk A. 1999. Acaulospora mellea and A. trappei, fungi new to the Mycota of Poland. Acta Mycol. 34, 41-50.
Dodd J. C., Arias I., Koomen I., Hayman D. S. 1990. The management of populations of vesicular-arbuscular mycorrhizal fungi in acid-infertile soils of a savanna ecosystem. Plant and Soil 122, 241-247.
Douds D. D., Schenck N. C. 1990. Relationship of colonization and sporulation by VA mycorrhizal fungi to plant nutrient and carbohydrate contents. New Phytol. 116, 621-627.
Estrada-Torres A., Varela L., Hernandez-Cuevas L., Cavito M. 1992. Some arbuscular mycorrhizal fungi from the state of Tlaxa. Mexico. Rev. Mex. Mic. 8, 85-110.
Grandi R. A. P., Trufem S. F. B. 1991. Fungos micorrízocos vesículo-arbusculares em Marantaceae cultivadas no Instituto de Botânica, São Paulo, SP. Revista brasil. Bot. 14, 89-95.
Koske R. E., Gemma J. N. 1997. Mycorrhizae and succession in plantings of beachgrass in sand dunes. Amer. J. Bot. 84, 118-130.
Mei-ging Z., You-shan W. 1992. Eight species of VA mycorrhizal fungi from northern China. Acta Mycol. Sinica 11, 258-267.
Morton J. B. 2000. International Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi. West Virginia University.
Musoko M., Last F. T., Mason P. A. 1994. Populations of spores of vesicular-arbuscular mycorrhizal fungi in undisturbed soils of secondary semideciduous moist tropical forest in Cameroon. Forest Ecol. Management 63, 359-377.
Saif S. R. 1987. Growth responses of tropical forage plant species to vesicular-arbuscular mycorrhizae. Plant and Soil 97, 25-35.
Schenck N. C., Spain J. L., Howeler R. H. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76, 685-699.
Sieverding E. 1989. Ecology of VAM fungi in tropical agrosystems. Agric. Ecosyst. Environ. 29, 369-390.
Sieverding E., Toro S. T. 1988. Influence of soil water regimes on VA mycorrhiza. V. J. Agr. Crop Sci. 161, 322-332.
Sieverding E., El-Sharkawy M., Hermandez A. P., Toro T. 1986. Influence of water stress on growth and formation of VA mycorrhiza of 20 cassava cultivars. In: Mycorrhizae: physiology and genetics. ESM, Dijon, 1-5 july 1985, INRA, Paris, 717-720.
Sylvia D. M. 1986. Spatial and temporal distribution of vesicular-arbuscular mycorrhizal fungi associated with Uniola paniculata in Florida foredunes. Mycologia 78, 728-734.