


In PVLG
Spain & N.C. Schenck
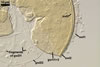
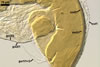
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germination walls.
|
|
|
In PVLG |
||
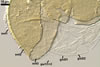
Layer 1 slowly deteriorating with age, hyaline, smooth, (0.7-)0.9(-1.2) µm thick when not degraded, folding or ballooning in spores crushed in lactic acid-based mountants, continuous with the wall of the sporiferous saccule. This layer usually is present in even older field-collected spores.
Layer 2 laminate, pale yellow (3A3) to yellow (3A6), (2.4-)3.1(-3.8) µm thick.
Layer 3 semiflexible, hyaline, 0.8-1.1 µm thick, tightly adherent to layer 2, separating from layer 2 only in vigorously crushed spores.
Germination wall 1 consists of two hyaline, semiflexible layers (gw1l1 and 2), each ca. 0.5 µm thick; these layers rarely separate from each other in crushed spores and, hence, are difficult to see.
 |
 |
 |
 |
 |
 |
 |
In PVLG |
||||||
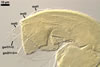
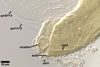
Germination wall 2 composed of two adherent layers (gw2l1 and 2).
Layer 1 flexible, hyaline, 0.5-0.8 µm thick, covered with small, <0.5 µm diam, granules usually scattering in crushed spores.
Layer 2 plastic, hyaline, 10-12 µm thick in PVLG, (0.5-)0.8(-1.5) µm thick and dark red (10C8-11C8) in Melzer’s reagent.
GERMINATION ORB. Not found.
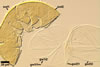
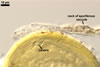
SPORIFEROUS SACCULE hyaline; globose to subglobose; 70-120 µm diam, with 1-layered wall, 0.7-1.2 µm thick; neck 70-110 µm long, 10.0-16.0 µm wide at the saccule, tapering to 6.0-8.0 µm wide at the spore attachment. Saccule usually collapses or falls off in mature spores.
 |
 |
In PVLG |
|
CICATRIX. A circular to ellipsoid scar, 5-9.5 x 5.5-13.0 µm.
MYCORRHIZAE. All the spores presented above were isolated from a field-collected rhizosphere soil of Calamagrostis arundinacea (L.) Roth. (Blaszkowski 1990). According to Morton (2002), the mycorrhizae of Ac. morrowiae consists of arbuscules, vesicles, and intraradical hyphae staining with considerably variation in intensity.
DISTRIBUTION. In Poland, Blaszkowski (1990) first recovered Ac. morrowiae (as Ac. rugosa) from among roots of C. rundinacea growing in a forest located in the Western Pomerania. This species has later been found to be one of the most frequently occurring arbuscular mycorrhizal fungus in the root zone of plants colonizing maritime dune soils of the Slowinski National Park (54º45'N, 17º26'E; Tadych and Blaszkowski 2000a), has been a frequent inhabitant of the inland dunes of the Bledowska Desert (50º22'N, 19º34'E; Blaszkowski et al. 2002), and has infrequently occurred in soils of the Tuchola Forests (53º46’N, 17º42’E-53º40’N, 17º54’E; Tadych and Blaszkowski 2000b).
Acaulospora morrowiae probably has a worldwide distribution, although its occurrence is rare. This fungus and Ac. longula (=Ac. morrowiae; see below) have been originally described from spores recovered from under native grasses growing in Carimagua, Colombia (Schenck et al. 1984). Morton (1986) found spores of Ac. morrowiae (as Ac. rugosa) in high aluminium, low pH soils of West Virginia.
PHYLOGENETIC POSITION. Unknown. However, if A. morrowiae is conspecific with A. longula and A. rugosa, as Morton (2002) suggested, the closest molecular relative of the former species is A. spinosa (Schwarzott et al. 2001).
NOTES. The holotype of A. morrowiae has been selected from spores produced in a one-species culture with Pueraria phaseoloides as the host plant (Schenck et al. 1984). This culture was established from spores originally isolated from under a sward of native grasses at Carimagua, Colombia.
The distinctive morphological characters of A. morrowiae are its light-coloured, yellow, small, and smooth spores with the outermost layer of their wall folding or ballooning when immersed in lactic acid-based mountants. In contrast to most the other species of the genus Acaulospora, in A. morrowiae this layer is relatively long-lived and was present in almost all spores isolated from field-collected soil samples by the author of this website. When intact, this layer resembles flexible, smooth layers of inner germinal walls of spores of the genera of, e. g., Acaulospora and Scutellospora.
Other species of the genus Acaulospora producing spores that may be easily confused with those of A. morrowiae because of their similarity in colour and size are A. delicata, A. gedanensis, and A. mellea.
At maturity, spores of A. morrowiae are pale yellow (3A3) to yellow (3A6), whereas mature spores of A. delicata are hyaline to pale yellow (3A3) and frequently have a green tint (Blaszkowski, pers. observ.; Morton 2002; Walker et al. 1986), which does not occur in spores of the former species.
The spore wall and the first inner germinal wall of spores of A. morrowiae consist of three and two layers, respectively, and not of two and one layer(s), respectively, as in spores of A. delicata (Blaszkowski, pers. observ.; Morton 2002). However, according to Morton (2002), the first germinal wall of the latter species may be 2-layered, similarly as in most other Acaulospora spp., but only one layer is visible.
Pale-coloured spores of A. gedanensis are identical to pale-coloured spores of A. morrowiae [pale yellow (3A3)], but the darkest spore-isolates of the former species [lemon yellow (3B8)] are much darker-coloured than the darkest specimens of the latter fungus [yellow (3A6)].
Spores of A. gedanensis generally are somewhat smaller than those of A. morrowiae [(55-)65(-75) µm diam when globose vs. (60-70-)76-92(-120) µm; Blaszkowski 1988; Schenck et al. 1984; Morton 2002].
Of the fungi compared here, A. morrowiae is most closely related morphologically to A. mellea. The fungi form spores of a similar size, as well as their subcellular structure and the phenotypic and biochemical properties of components of the spore wall and the two inner germinal walls are almost identical. Spores of A. mellea may be darker-coloured [pale yellow (3A3) to orange (5B8) vs. pale yellow (3A3) to yellow (3A6) in A. morrowiae] and their outermost spore wall layer usually quickly sloughs and never balloons in lactic acid-based mountants (vs. it slowly deteriorates with age and highly swells in lactic acid-based mountants; Blaszkowski et al. 1999; Morton 2002).
According to Morton (2002), A. morrowiae is synonymous with A. rugosa and A. longula.
As mentioned above (see Phylogenetic position), the closest molecular relative of A. morrowiae probably is A. spinosa. However, morphologically these fungi are easy to distinguish. The most evident difference readily separating them resides in the structural laminate wall layer of their spores. While the upper surface of the laminate spore wall layer of A. morrowiae is smooth, that of A. spinosa is ornamented with rounded spines, 2-3 µm long when seen in a cross view (Morton 2002; Walker and Trappe 1981).
REFERENCES
Blaszkowski J. 1988. Four new species of the Endogonaceae (Zygomycotina) from Poland. Karstenia 27, 37-42.
Blaszkowski J. 1990. Polish Endogonaceae. II. Acaulospora rugosa, Glomus aggregatum, Glomus etunicatum, Glomus fasciculatum and Glomus occultum. Karstenia 30, 1-13.
Blaszkowski J., Tadych M., Madej T. 2002. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of the Bledowska Desert, Poland. Acta Soc. Bot. Pol. 71, 71-85.
Blaszkowski J., Tadych M., Madej T., Adamska I., Czerniawska B., Iwaniuk A. 1999. Acaulospora mellea and A. trappei, fungi new to the Mycota of Poland. Acta Mycol. 34, 41-50.
Morton J. B. 1986. Three new species of Acaulospora (Endogonaceae) from high aluminum, low pH soils in West Virginia. Mycologia 78, 641-648.
Morton J. B. 2002. International Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi. West Virginia University. http://www.invam.caf.wvu.edu/.
Schenck N. C., Spain J. L., Howeler R. H. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76, 685-699.
Schwarzott D., Walker C., Schüßler A. 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales) is nonmonophyletic. Mol. Phyl. Evol. 21, 190-197.
Tadych M., Blaszkowski J. 2000a. Arbuscular fungi and mycorrhizae (Glomales) of the Slowinski National Park, Poland. Mycotaxon 74, 463-483.
Tadych M., Blaszkowski J. 2000b. Arbuscular mycorrhizal fungi of the Brda river valley in the Tuchola Forests. Acta Mycol. 35, 3-23.
Walker C., Pfeiffer C. M., Bloss H. E. 1986. Acaulospora delicata sp. nov. an endomycorrhizal fungus from Arizona. Mycotaxon 25, 621-628.
Walker C., Trappe J. M. 1981. Acaulospora spinosa sp. nov. with a key to the species of Acaulospora. Mycotaxon 12, 515-521.