

In PVLG
|
|
|
|
|
In PVLG |
||||
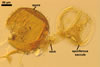
SPORES single in the soil or in aggregates without a peridium. Aggregates 160-520 x 300-700 µm, composed of 2-5 spores. Spores develop laterally on the neck of a sporiferous saccule; brownish orange (6C8) to brown (6E8); globose to subglobose; (150-)185(-240) µm diam.
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germinal walls.
Spore wall composed of three layers (swl1-3).
 |
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's reagent |
||||
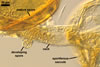
Layer 1 semipermanent, hyaline, (2.5-)4.4(-6.5) µm thick, continuous with the wall of the sporiferous saccule, usually intact in mature spores.
Layer 2 laminate, smooth, brownish orange (6C8) to brown (6E8), (2.5-)3.3(-4.0) µm thick.
Layer 3 semiflexible, hyaline, (1.3-)1.5(-1.8) µm thick, easily separating from layer 2.
Germinal wall 1 composed of two hyaline, tightly adherent layers (gw1l1 and 2), each ca. 0.5 µm thick.
 |
 |
 |
 |
 |
In PVLG+Melzer's reagent |
||||
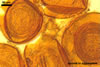
Layer 1 flexible, ca. 0.5 µm thick, ornamented with small, <0.5 µm diam, granules.
Layer 2 plastic, 10-15 µm thick in PVLG, (0.5-)0.9(-1.0) µm thick and staining beetroot purple (13D8) in Melzer’s reagent.
GERMINATION ORB. Not found.
 |
In PVLG |
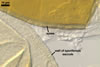
SPORIFEROUS SACCULE hyaline to pale yellow (3A3); globose to subglobose; 170-210 µm diam; neck 70-250 µm long, 35-45 µm wide at the saccule, tapering to 20-38 µm wide at the spore attachment. Saccule usually collapses or falls off in mature spores.
 |
In PVLG |
CICATRIX. A slightly raised collar when seen in a cross view, circular, 11.5-17.0 µm diam, or ellipsoidal, 11.5-13.0 x 12.0-17.0 µm when observed in a plane view.
MYCORRHIZAE. Acaulospora thomii has been associated in the field with vesicular-arbuscular roots of Triticum aestivum L. Many attempts to establish mycorrhizae of this fungus in one-species cultures with Plantago lanceolata L. failed.
DISTRIBUTION. Acaulospora thomii has been described based on spores isolated from under T. aestivum cultivated in Boleslawice (51o18’N, 15º22’E) in south-western Poland (Blaszkowski 1988). Dr. F. Oehl (pers. comm.) found spores of this fungus in cultivated soils of Switzerland.
NOTES. Acaulospora thomii distinguishes its brown-coloured and relatively large spores formed both singly and in loose aggregates.
The species of the genus Acaulospora producing brown-coloured spores of a similar size range to that of spores of A. thomii are only A. sporocarpia and A. walkeri. Moreover, A. sporocarpia also forms spores both singly and in aggregates. However, spores of A. sporocarpia may be much darker-coloured (dark brown to black; Berch 1985), and the outermost wall layer of spores of A. walkeri may be pale yellow to yellow brown (Kramadibrata and Hedger 1990), and not hyaline as that of spores of A. thomii. Both the former species probably should be redescribed, because both the subcellular structure of their spores and the phenotypic and biochemical properties of its components do not fit those occurring in spores of properly defined Acaulospora spp.
Spores of A. capsicula, A. koskei, and A. laevis overlap in size with those of A. thomii and, except for A. koskei, have an identical subcellular structure with its components of the same phenotypic and biochemical properties (Blaszkowski 1988, 1990, 1995, 2003; Morton 2002). In the first germinal wall of spores of A. koskei, only one layer is visible (vs. two layers in A. homii), although this wall may comprise two tightly adherent, and thereby difficult to detect, layers, as Morton (2002) concluded. However, compared with the brownish orange (6C8) to brown (6E8) spores of A. thomii occurring singly and in loose aggregates, those of A. capsicula are orange red (8A8) to capsicum red (8B8), A. koskei orange white (5A2) to reddish orange (7A8), A. laevis salmon to orange, and origin only singly in the soil (Blaszkowski 1988, 1990, 1995, 2003, 2003; Morton 2002). Finally, the outermost spore wall layer of the three latter species usually is completely sloughed in mature specimens, whereas it was present in most field-collected spores of the former fungus (Blaszkowski 1988).
Coloured spores of the other known species of the genus Acaulospora of a smooth upper surface of their laminate spore wall layer are either markedly smaller (A. mellea, A. morrowiae; Blaszkowski 2003; Morton 2002; Schenck et al. 1984) or larger (A. entreira; Velazquez et al. 2008) than those of the fungus discussed here.
REFERENCES
Berch S. M. 1985. Acaulospora sporocarpia, a new sporocarpic species, and emendation of the genus Acaulospora (Endogonaceae, Zygomycotina). Mycotaxon 23, 409-418.
Blaszkowski J. 1988. Four new species of the Endogonaceae (Zygomycotina) from Poland. Karstenia 27, 37-42.
Blaszkowski J. 1990. Polish Endogonaceae. VII. Acaulospora capsicula sp. nov. Mycologia 82, 794-798.
Blaszkowski J. 1995. Acaulospora koskei, a new species in Glomales from Poland. Mycol. Res. 99, 237-240.
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland. http://www.agro.ar.szczecin.pl/~jblaszkowski/.
Kramadibrata K., Hedger J. N. 1990. A new species of Acaulospora associated with cocoa in Java and Bali (Indonesia). Mycotaxon 37, 73-77.
Morton J. B. 2002. International Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi. West Virginia University: http://www.invam.caf.wvu.edu/.
Schenck N. C., Spain J. L., Howeler R. H. 1984. Several new and unreported vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Colombia. Mycologia 76, 685-699.
Velazquez M. S., Cabello M., Irrazabal G., Godeas A. 2008. Acaulosporaceae from El Palmar National Park, Entre Rios, Argentina. Mycotaxon 103, 171-187.