(Gerd. & B.K. Bakshi) R.T. Almeida & N.C. Schenck
 |
 |
 |
 |
In PVLG |
|||
(Gerd. & B.K. Bakshi) R.T. Almeida & N.C. Schenck
|
|
|
|
In PVLG |
|||
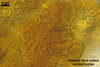
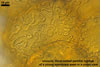
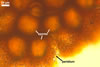
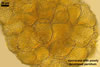
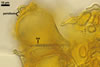
Sporocarps orange (5B8) to brownish orange (6C8); globose to subglobose; (250-)303(-380) µm diam; to ellipsoid; 300-420 x 510-560 µm; frequently tuberculate from protruding spores; with a peridium and two to six monohyphal stalks.
Peridium 5.3-25.5 µm thick, composed of sinuous, septate hyphae, (2.7-)4.5(-7.1) µm wide, of a wall of (1.0-)1.5(-2.0) µm thick, tightly enclosing a group of spores. The peridium seems to develop from the monohyphal stalks (Wu 1993) and is abundant in even immature spores (Morton 2002).
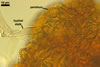
Monohyphal stalk hyaline to yellowish white (4A2), straight or slightly curved, septate, occasionally branched, ca. 5.0-8.0 µm wide, with a wall ca. 1.0-1.5 µm thick.
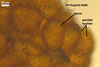
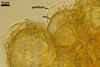
Spores pale yellow (4A3) to reddish yellow (4A7); pulvinate, elliptical to clavate; 28-62 x 58-88 µm; more rarely globose to subglobose; (62-)64(-80) µm diam; radiating side by side in a single layer from a central plexus of hyphae; with one subtending hypha.
SUBCELLULAR STRUCTURE OF SPORES consists of a one-layered, smooth, laminate, pale yellow (4A3) to reddish yellow (4A7), (1.7-)2.7(-3.2) µm thick layer (swl1).
 |
 |
 |
 |
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's |
|||||||
 |
 |
In PVLG |
|
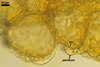
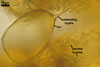
SUBTENDING HYPHA straight or recurved; cylindrical to slightly flared; (4.4-)5.9(-7.4) µm wide at the spore base.
Wall of subtending hypha pale yellow (4A3) to reddish yellow (4A7); (2.0-)2.5(-2.9) µm thick at the spore base; composed of one layer continuous with the spore wall.Pore (1.5-)2.0(-2.5) µm diam, open or occluded by a curved septum continuous with the innermost laminae of the spore wall.
 |
 |
In PVLG |
|
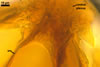
CENTRAL PLEXUS 60-250(-340) µm diam, composed of tightly interwoven, hyaline to yellowish white (4A2), septate, thin-walled hyphae, ca. 5.0-5.8 µm wide.
Neither spores nor peridial and central plexus hyphae stain in Melzer's reagent.
GERMINATION. By germ hyphae developing from the central hyphal plexus (Almeida and Schenck 1990). The germ hyphae produce extramatrical, vesicle-like, mostly globose to subglobose structures, up to 35 µm diam, which form terminally and intercalary.
MYCORRHIZAE. According to Almeida and Schenck (1990) and Morton (2002), Gl. sinuosum formed mycorrhizae with arbuscules, vesicles, as well as with extra- and intraradical hyphae staining intensively in trypan blue.
PHYLOGENETIC POSITION. Schwarzott et al. (2001) found Gl. sinuosum to be a member of the subclade b of Glomus group A also comprising Gl. coremioides (Berk. & Broome) D. Redecker & J.B. Morton, Gl. clarum Nicol. & N.C. Schenck, Gl. fasciculatum (Thaxt.) Gerd. & Trappe emend. C. Walker & Koske, Gl. intraradices N.C. Schenck & G.S. Sm., Gl. proliferum Dalpe & Declerck, and Gl. vesiculiferum (Thaxt.) Gerd. & Trappe.
DISTRIBUTION. The type of Gl. sinuosum has been isolated from under Fraxinus uhdei (Wenz) Lingelsh. growing in the Demonstration Forest, F. R. I., located in Dehra Dun, Uttar Pradesh, India (Gerdemann and Bakshi 1976). The date of the collection was September 1974. Additionally, the same scientists found sporocarps of this fungus in rhizosphere soils of many other plant species.
Other literature reports of the finding of Gl. sinuosum in different cultivated and uncultivated sites are, e. g., those from USA (Haas and Menge 1990; Miller et al. 1985; Schenck and Kinloch 1980; Menge et al. 1977; Nicolson and Schenck 1979), Hawaii (Koske 1988; Koske and Gemma 1996), Mexico (Estrada-Torres et al. 1992; Guadarrama and Alvarez-Sanchez 1999), Colombia (Sieverding 1989), Brazil (Moreira et al. 2007), Switzerland (Oehl et al. 2005), Israel (Blaszkowski et al. 2001; Dodd and Krikun 1984; Haas and Menge 1990), China (Gai et al. 2006), and Pakistan (Iqbal and Perveen 1980).NOTES. Glomus sinuosum presented above was characterized based on its specimens (slides ......) obtained from Dr. F. Oehl, .........., those extracted from field samples collected by ....harikumar...... from under ....... growing ........., Africa, and descriptions of this fungus made by Almeida and Schenck (1990), Morton (2002) and Wu (1993).
Only a few properties of the specimens of Gl. sinuosum determined by me more or less diverged from those published by Almeida and Schenck (1990), Gerdemann and Bakshi (1976), Morton (2002), and Wu (1993). First, compared with Morton's (2002) description, the wall of peridial hyphae and that of spores of Gl. sinuosum examined by me were thicker and thinner, respectively [0.5-1.0 µm thick and 1.5-6.0 µm thick, respectively vs. (1.0)1.5(-2.0) µm thick and (1.7-)2.7(-3.2) µm thick, respectively]. The range of thickness of the spore wall given by Almeida and Schenck (1990), Gerdemann and Bakshi (1976), and Wu (1993) also was slightly wider than that determined by the author of this website (1.3-4.9 µm thick vs. 1.7-3.2 µm thick). Second, as results from the descriptions of Almeida and Schenck (1990) and Wu (1993), sporocarps and spores of Gl. sinuosum may be much larger (up to 650 µm and 45-150 x 30-83 µm, respectively) than those characterized here (up to 560 µm and 28-62 x 58-88 µm, respectively).
The unique structure of Gl. sinuosum is its peridium consisting of sinuous, thick-walled hyphae forming a compact covering enclosing a group of spores. To extract the spores, the peridium has to be more or less physically removed.
Of the known Glomus spp. forming spores in sporocarps with a peridium, the fungi most resembling Gl. sinuosum morphologically are Gl. coremioides, Gl. fuegianum (Speg.) Trappe & Gerd., and Gl. liquidambaris (C.G. Wu & Z.C. Chen) R.T. Almeida & N.C. Schenck.
Looking at intact sporocarps under a compound microscope, first marked differences between the species listed above are visible in the morphology of their peridium. As mentioned above, the peridium of Gl. sinuosum sporocarps consists of sinuous, thick-walled hyphae forming a compact covering enclosing a group of spores. In contrast, in Gl. coremioides, the peridium is composed of interwoven hyphae (Almeida and Schenck 1990; Wu 1993), and not sinuous ones. Sporocarps of Gl. fuegianum may lack a peridium, which, when present, consists of thin-walled, also interwoven hyphae (Blaszkowski 2002; Blaszkowski et al. 1998). In Gl. sinuosum, the peridium is always formed. The peridium of Gl. liquidambaris sporocarps is formed by cylindrical, clavate, or spathulate paraphysis-like structures (Almeida and Schenck 1990; Wu 1993). They develop from either the central hyphal plexus of the sporocarps or the base of their spores and protrude over the upper wall of the radially distributed spores.
Additionally, compared with Gl. sinuosum producing sporocarps singly or in groups of two, Gl. coremioides forms conglomerates consisting of many sporocarps fused together laterally or vertically forming columns up to four sporocarps (Almeida and Schenck 1990; Wu 1993). Moreover, individual or groups of Gl. coremioides sporocarps sometimes are covered with whitish to reddish crust, of the upper surface formed by thin-walled, vesicle-like structures, which are lacking in Gl. sinuosum. Finally, in contrast to the nonreactivity of any structure of Gl. sinuosum sporocarps in Melzer's reagent (Almeida and Schenck 1990; Blaszkowski, pers. observ.), hyphae of peridium, plexus and a short stalk of Gl. coremioides sporocarps stain blue in this reagent (Almeida and Schenck 1990).
Apart from the differences characterized above, four other morphological properties separate Gl. sinuosum and Gl. fuegianum. First, spores of Gl. fuegianum form in groups or sporocarps comprising from 2 to 28 sporocarps, each consisting of 3 to 13 radially distributed spores developed from a thick-walled, inflated hypha (Blaszkowski 2002; Blaszkowski et al. 1998). In contrast, spores of Gl. sinuosum occur only in sporocarps and origin at the tips of tightly interwoven, thin-walled hyphae of the central plexus (Almeida and Schenck 1990; Blaszkowski, pers. observ.; Wu 1993). Second, single spores of Gl. fuegianum are sometimes surrounded with branched and convoluted hyphae, a phenomenon not occurring in Gl. sinuosum. Third, Gl. fuegianum sporocarps are of a yellow to brown tint, and not orange to brownish orange as those of Gl. sinuosum. Forth, the spore wall of Gl. fuegianum consists of two layers, whereas that of Gl. sinuosum is one-layered, lacking the evanescent layer forming the spore surface of the former fungus.
Glomus liquidambaris and Gl. sinuosum are similar in size and colour of their sporocarps, as well as in size of their spores (Almeida and Schenck 1990; Blaszkowski, pers. observ.; Morton 2002; Wu 1993). However, most spores of the former species are clavate and have a highly thickened top wall, whereas spores of the latter fungus, although may be also clavate (Almeida and Schenck 1990; Wu 1993), generally are globose to egg-shaped and their wall is more or less even in thickness (Blaszkowski, pers. observ.).
Apart from Gl. coremioides, the only species of a recognized phylogenetic position of the fungi compared above, results of Schwarzott's et al. (2001) investigations showed that other members of the Glomeromycota, of so far determined molecular properties, closely related to Gl. sinuosum also are Gl. clarum, Gl. intraradices, Gl. fasciculatum, Gl. proliferum, and Gl. vesiculiferum. However, of them, only Gl. vesiculiferum produces sporocarps with a peridium. Three morphological characters easily separate Gl. sinuosum from Gl. vesiculiferum. First, sporocarps of Gl. sinuosum are darker coloured and much smaller [orange (5B8) to brownish orange (6C8) and (250-)303(-380) µm diam, respectively; Blaszkowski, pers. observ.] than those of Gl. vesiculiferum (whitish and 13x10 mm, respectively; Gerdemann and Trappe 1974). Second, while the peridium of Gl. sinuosum sporocarps consists of sinuous, thick-walled hyphae (Almeida and Schenck 1990; Blaszkowski, pers. observ.; Wu 1993), thin-walled, globose to broadly clavate vesicles form the peridium of Gl. vesiculiferum sporocarps (Gerdemann and Trappe 1974). Third, in contrast to the one-layered, laminate spore wall of Gl. sinuosum (Almeida and Schenck 1990; Blaszkowski, pers. observ.; Wu 1993), the wall of Gl. vesiculiferum spores consists of two layers: an evanescent outer layer and a laminate inner one (Gerdemann and Trappe 1974).
Glomus sinuosum has originally been described as Sclerocystis sinuosa Gerd. & Bakshi (Gerdemann and Bakshi 1976). Considering differences in the mode of sporocarp formation and spore development, Almeida and Schenck (1990) remained in the genus Sclerocystis Berk. & Broome emend. R.T. Almeida & N.C. Schenck only S. coremioides Berk. & Broome. Of the 13 other described Sclerocystis spp., five were transferred to the genus Glomus Tul. & C. Tul. [as Gl. clavisporum, Gl. liquidambaris (C.G. Wu & Z.C. Chen) R.T. Almeida & N.C. Schenck, Gl. rubiforme (Gerd. & Trappe) R.T. Almeida & N.C. Schenck, Gl. sinuosum, Gl. taiwanense (C.G. Wu & Z.C. Chen) R.T. Almeida & N.C. Schenck], and eight were considered to be synonymous. Wu (1993b) rejected the changes of Almeida and Schenck (1990) and reverted to the previous classification of Gerdemann and Trappe (1974). However, phylogenetic analyses of the 18S ribosomal subunit of S. coremioides and S. sinuosa have showed them to be members of a monophyletic clade comprising, e. g., Gl. intraradices N.C. Schenck & G.S. Sm. and Gl. mosseae (Nicol. & Gerd.) Gerd. & Trappe (Redecker et al. 2000). Consequently, S. coremioides was renamed Gl. coremioides and the genus Sclerocystis stopped to exist.
REFERENCES
Almeida, R. T., and N. C. Schenck. 1990. A revision of the genus Sclerocystis (Glomaceae, Glomales). Mycologia 82, 703-714.
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone, and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland.http://www.agro.ar.szczecin.pl/~jblaszkowski/.
Blaszkowski J., Madej T., Tadych M. 1998. Entrophospora baltica sp. nov. and Glomus fuegianum, two species in the Glomales from Poland . Mycotaxon 68, 165-184.
Blaszkowski J., Tadych M., Madej T., Adamska I., Iwaniuk A. 2001. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of Israeli soils. Mat. II Polsko-Izraelskiej Konf. Nauk. nt. "Gospodarowanie zasobami wodnymi i nawadnianie roslin uprawnych". Przeglad naukowy Wydz. Inz. Ksztalt. Srod. 22, 8-27.
Dodd J. C., Krikun J. 1984. Observations on endogonaceous spores in the Negev Desert. Trans. Br. Mycol. Soc. 82, 536-540.
Estrada-Torres A., Varela L., Hernandez-Cuevas L., Cavito M. E. 1992. Algunos hongos micorrizicos arbusculares del estado de Tlaxcala, México. Rev. Mex. Mic. 8, 85-110.
Gai J. P., Christie P., Feng G., Li X. L. 2006. Twenty years of research on biodiversity and distribution of arbuscular mycorrhizal fungi in China : a review. Mycorrhiza 16, 229-239.
Gerdemann J. W., Bakshi B. K. 1976. Endogonaceae of India: two new species. Trans. Br. Mycol. Soc. 66, 340-343.
Gerdemann J. W., Trappe J. M. 1974. The Endogonaceae in the Pacific Northwest. Myc. Memoir 5, 1-76.
Guadarrama P., Alvarez-Sanchez F. J. 1999. Abundance of arbuscular mycorrhizal fungi spores in different environments in a tropical rain forest, Veracruz, Mexico. Mycorrhiza 8, 267-270.
Haas J. H., Menge J. A. 1990. VA-mycorrhizal fungi and soil characteristics in avocado (Persea americana Mill.) orchard soils. Plant and Soil 127, 207-212.
Iqbal S. H., Perveen B. 1980. Some species of Sclerocystis (Endogonaceae) from Pakistan. Trans. Mycol. Soc. Japan 21, 57-63.
Koske R. E. 1988. Vesicular-arbuscular mycorrhizae of some Hawaiian dune plants. Pac. Sci. 42, 217-229.
Koske R. E., Gemma J. N. 1996. Arbuscular mycorrhizal fungi in Hawaiian sand dunes: Island of Kaua'i. Pacific Sci. 50, 36-45.
Menge J. A., Nemec S., Davis R. M., Minassian V. 1977. Mycorrhizal fungi associated with citrus and their possible interactions with pathogens. Proc. Int. Soc. Citriculture 3, 872-876.
Miller D. D., Domoto P., Walker C. 1985. Mycorrhizal fungi at eighteen apple rootstock plantings in the United States. New Phytol. 100, 379-391.
Moreira M., Nogueira M. A., Tsai S. M., Gomes-da-Costa S. M., Cardoso E. J. B. N. 2007. Sporulation and diversity of arbuscular mycorrhizal fungi in Brazil Pine in the field and in the greenhouse. Mycorrhiza 17, 519-526.
Morton J. B. 2002. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University.http://www.invam.caf.wvu.edu/.
Nicolson T. H., Schenck N. C. 1979. Endogonaceous mycorrhizal endophytes in Florida. Mycologia 71, 178-198.
Oehl F., Sieverding E., Ineichen K., Ris E.-A., Boller T., Wiemken A. 2005. Community structure of arbuscular mycorrhizal fungi at different soil depths in extensively and intensively managed agroecosystems. New Phytol. 165, 273-283.
Redecker D., Morton J. B., Bruns T. D. 2000. Molecular phylogeny of the arbuscular mycorrhizal fungi Glomus sinuosum and Sclerocystis coremioides. Mycologia 92, 282-285.
Schenck N. C., Kinloch R. A. 1980. Incidence of mycorrhizal ungi on six field crops in monoculture on a newly cleared woodland site. Mycologia 72, 445-456.
Schwarzott D., Walker C., Schüßler A. 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales) is nonmonophyletic. Mol. Phyl. Evol. 21, 190-197.
Sieverding E. 1989. Ecology of VAM fungi in tropical ecosystems. Agric., Ecosyst., Environ. 29, 369-390.
Wu C.-G. 1993. Glomales of Taiwan: IV. A monograph of Sclerocystis (Glomaceae). Mycotaxon 49, 327-349.