|
|
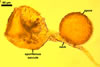
SPORES formed singly in the soil; originating blastically at the tip of a hyphal branch (pedicel) of the neck of a sporiferous saccule. Spores pale yellow (4A3) to maize yellow (4A6); globose to subglobose; (150-)210(-250) µm diam; sometimes ovoid; 150-190 x 210-250 µm.
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's |
|||
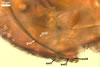
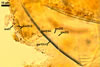
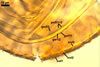
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germinal walls.
Spore wall composed of three layers (swl1-3).
 |
 |
 |
 |
 |
 |
 |
 |
 |
In PVLG |
||||||||
 |
 |
 |
In PVLG+Melzer's reagent |
||
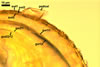
Layer 1, forming the spore surface, evanescent, pale yellow (4A3) to maize yellow (4A6), (1.7-)3.3(-5.6) µm thick when intact, usually with deep fissures due to sloughing with age.
Layer 2 semi-flexible, smooth, hyaline (1.2-)2.5(-3.7) µm thick, continuous with the wall of the pedicel.
Layer 3 flexible to semi-flexible, hyaline, 0.5-1.0 µm thick, usually tightly adherent to the lower surface of layer 2 and, hence, difficult to see.
Germinal wall 1 comprises two fragile, smooth, hyaline layers (gw1l1 and 2), (0.5-)1.1(-1.6) µm and (0.6-)1.4(-2.1) µm thick, respectively, usually tightly adherent to one another and readily cracking to form line slits or polygonal pieces in crushed spores.
Germinal wall 2 consists of three smooth, hyaline layers (gw2l1-3).
 |
 |
In PVLG |
|
Layer 1 flexible to semi-flexible, 0.5-0.8 µm thick, always tightly adherent to layer 2 and, thereby, very difficult to observe.
Layer 2 semi-flexible, finely laminate, hyaline, (1.7-)2.8(-4.1) µm thick.
Layer 3 flexible to semi-flexible, 0.5-0.8 µm thick, rarely separating from layer 2.
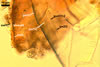
In Melzer's reagent, only spore wall layer 1 stains orange (6A6) to pastel red (8C5).
|
In PVLG |
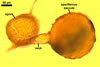
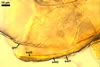
PEDICEL 10-15 µm long, 15-27 µm wide at the spore base, positioned 80-120 µm from the base of the sporiferous saccule, consisting of a hyaline wall continuous with the wall of the saccule neck, the spore wall, and layer 1 of germinal wall 1. Pore open. According to Spain et al. (2006), in young spores, the pore is closed by germinal wall 1.
GERMINATION ORB. Not found.
SPORIFEROUS SACCULE hyaline to yellowish white (4A2), globose to subglobose, (185-)200(-275) µm diam, occasionally ovoid, 160-200 x 210-260 µm, formed at the end of a funnel-shaped neck.
Wall of sporiferous saccule semi-flexible, semi-permanent, hyaline to yellowish white (4A2), 3.5-8.8 µm thick, smooth in young specimens, roughened because of a patchy sloughing of its upper surface, composed of two to three tightly adherent layers, usually difficult to observe.
Saccule neck hyaline to yellowish white (4A2), 200-250 µm long, 30-45 µm wide at the base of the saccule, 20-25 µm wide at the spore base, then gradually tapering up to 8-10 µm wide.
 |
In PVLG |
CICATRIX. A slightly raised collar when seen in a cross view, circular, 9.5-12.0 µm diam, or ellipsoidal, 9.5-12.0 x 12.0-14.0 µm when observed in a plane view.
MYCORRHIZAE. In the field, spores of Am. gerdemannii occurred among vesicular-arbuscular mycorrhizal roots of Achillea millefolium L., Campanula polymorpha Witasek, Cardaminopsis neglecta (Schult) Hayek, Cerastium tatrae Borbás, Equisetum arvense L., Hypericum perforatum L., Juniperus communis L., Knautia kitaibelii (Schult.) Borbás, Leucanthemum waldsteinii (Sch.Bip.) Pouzar, Melamphyrum herbichii Wol., Sesleria tatrae (Degen) Deyl, Thuja occidentalis L., and Thymus sp. Additionally, they were recovered from some trap cultures established from mixtures of roots and rhizosphere soils of these plants and with P. lanceolata as the plant host. Unfortunately, many attempts to produce one-species cultures of Am. gerdemannii failed.
According to Morton (2002) and Morton and Redecker (2001), mycorrhizae of Am. gerdemannii consisted of only arbuscules and intraradical hyphae; no vesicles were found. All the structures were patchily distributed along roots and stained weakly or not at all in trypan blue. Percentage mycorrhizal colonization always was very low, below 10%.
DISTRIBUTION. In Poland, spores of Am. gerdemannii were found in 19 samples of roots and rhizosphere soils of 14 species of uncultivated plants. None of the ca. 2000 root and soil mixtures coming from cultivated sites of Poland contained spores of this fungus.
The average abundance of spores of Am. gerdemannii in the samples examined was 4.7 and ranged from 1 to 29 in 100 g dry soil. The proportion of spores of this species in spore populations of all the arbuscular fungi recovered averaged 11.8% in a range of 2.1-66.7%. The average abundance of species of arbuscular fungi in samples in which spores of Am. gerdemannii occurred was 2.7 and ranged from 1 to 6 in 100 g dry soil.
The arbuscular fungi accompanying Am. gerdemannii in the field were Acaulospora bireticulata, A. capsicula, A. koskei, A. lacunosa, A. paulinae, Archaeospora trappei, Entrophospora infrequens, Gl. caledonium, Gl. claroideum, Gl. constrictum, Gl. deserticola, Gl. ? etunicatum, Gl. fasciculatum, Gl. geosporum, Gl. macrocarpum, Gl. mosseae, Gl. pansihalos, an unrecognized Glomus sp., Pacispora scintillans, Scutellospora dipurpurescens, and S. pellucida.
Although Am. gerdemannii probably has a worldwide distribution, this species has infrequently been recorded. Most reports of Am. gerdemannii come from the United States of America (Allen and MacMahon 1985; An et al. 1990, 1993a, b, c; Bever et al. 1996; Koske et al. 1977; Nicolson and Schenck 1979; Rose et al. 1979). Additionally, spores of this fungus have been isolated in Brazil (Moreira-Souza et al. 2003), Colombia (Dodd et al. 1990), and Australia (Morton and Redecker 2001). In southern Poland, Turnau et al. (2001) recovered spores of a morphotype named Glomus sp. HM-CL4 of molecular properties close (71%) to those of Gl. gerdemannii, the basionym of Am. gerdemannii (Spain et al. 2006). However, the authors did not show any morphological characters of the spores isolated and did not determine which of the two morphotypes of this fungus was found.
NOTES. The distinctive properties of Am. gerdemannii are its fragile bi-layered germinal wall 1 and the mode of differentiation of spores of this fungus. Additionally, Am. gerdemannii is a dimorphic fungus producing two morphotypes, acaulosporioid (as in Acaulospora spp.) and glomoid (as in Glomus spp.), a phenomenon rarely occurring in other species of arbuscular fungi.
The germinal wall 1 of Am. gerdemannii resembles a hardened glass. In slightly crushed spores, linear cracks appear on the surface of this wall. However, in spores vigorously crushed, this wall almost always breaks into small pieces. Another characteristic of this wall is its glittering when seen in a polarized light.
The author of this website was unable to produced one-species cultures of Am. gerdemannii, and spores of this fungus recovered from both field-collected samples and trap cultures did not make possible to investigate their whole ontogenetic development. According to Morton and Redecker (2001), the ontogenetic development of the acaulosporioid morphotype of Am. gerdemannii starts the formation of a sporiferous saccule at the end of the sporiferous saccule neck continuous with an extraradical hypha of an arbuscular mycorrhiza, identically to that of members of the genus Acaulospora. Spores of this fungus originate from a hyphal branch called a pedicel positioned 180-260 µm from the base of the saccule. The wall of the pedicel consists of hyaline layers continuous with layers of the spore wall and layer 1 of germinal wall 1. The pedicels of Polish specimens were much shorter (10-15 µm long) than those characterized by Morton and Redecker (2001; 50-90 µm long) and, thereby, undoubtedly formed a more persistent link between spores and sporiferous saccules than longer ones. All the specimens of Am. gerdemannii found by the authors of this paper came from the field and probably only the sporiferous saccules with short pedicels remained associated with spores during their isolation. Sporiferous saccules rarely occur in field-collected spores of the families Acaulosporaceae and Archaeosporaceae because of their detachment or deterioration by soil microorganisms (Blaszkowski 2003). When spores begin to expand blastically at the end of the pedicel, spore wall layer 1 forms de novo. In young spores, it is smooth and best visible in specimens mounted in Melzer's reagent in which it stains orange (6A6) to pastel red (8C5). However, with age, it becomes friable, degrades, and sloughs partly in separated patches or completely. Hence, older spores are either covered with yellow brown remnants of layer 1 or are hyaline because of the prominence of the second hyaline spore wall layer. After the full differentiation of the outermost layer 1 and the innermost layer 3 of the spore wall, first the 2-layered germinal wall 1 and then the 3-layered germinal wall 2 probably successively origin, similarly as in spores of members of the genus Acaulospora. In fully mature spores, the germinal wall 2 resembles a coriaceous wall sensu Walker (1986). However, by definition, a coriaceous wall is a uniform structure, whereas the germinal wall 2 of Am. gerdemannii consists of three layers, of which the middle one corresponds with properties of a laminate layer. Except for Ambispora spp., no other species of the phylum Glomeromycota forms spores with a laminate layer of their innermost wall. Although Morton and Redecker (2001) also found the germinal wall 1 (called layer 3) to be fragile and the germinal wall 2 (called layer 4) to resemble a skin, they did not detect their 2- and 3-layered structure, respectively. According to Morton and Redecker (2001), layers 3 and 4 are uniform structures. Additionally, they concluded that these layers are components of the spore wall. However, only the outer layer of the germinal wall 1 may arise from the pedicel wall with which layers of the spore wall are continuous (Spain et al., in press) and the germinal wall 1 has no physical contact with the innermost germinal wall 2. Moreover, all these walls usually markedly separate from each other in crushed spores. Thus, the location of these walls in relation to one another and to the spore wall highly resembles that of the spore walls and germinal walls of spores of fungi of the genera Acaulospora, Entrophospora, Kuklospora, Pacispora, and Scutellospora (Blaszkowski 2003). The outer surface of the innermost germinal wall of spores of the latter genera is the place on which pre-germination structures (called germination orb or germination shield) form with emerging germ tubes. Although no one of such structures were so far revealed in spores of Am. gerdemannii, the formation of the germinal wall 2 in spores of this species probably precedes the synthesis of a similar structure and, thereby, makes the fungus to be able to germinate.
When observed under a dissecting microscope, spores of Am. gerdemannii may easily be confused with those of A. spinosa, A. thomii, and Am. appendicula and Am. fennica. All the fungi form yellow brown and dull spores of a similar size range.
Examination of the subcellular structure of spores and their ontogenetic development readily separates the species of Ambispora from those of Acaulospora. Spores of Am. appendicula, Am. fennica, and Am. gerdemannii differ mainly in the properties of layers of the first inner germinal wall. In the latter two species, both layers of this wall are smooth on both sides. In contrast, in the former fungus, the lower surface of layer 1 is ornamented with hemispherical protuberances, which impress hemispherical concave depressions on the upper surface of layer 2 during its differentiation (Morton and Redecker 2001). Additionally, in vigorously crushed spores, the germinal wall 1 of Am. fennica and Am. gerdemannii usually disintegrates, and that of Am. appendicula remains its integrity. As concluded in the page characterizing Am. fennica, Am. fennica and Am. gerdemannii probably are congeneric.
The main differences between Am. gerdemannii, A. spinosa, and A. thomii are (1) the continuity of only the outermost spore wall layer with the wall of a sporiferous saccule in Acaulospora spp. (vs. the wall of a sporiferous saccule is continuous with spore wall layers 1-3 and layer 1 of germinal wall 1 in Am. gerdemannii) and (2) the presence of a beaded outer layer and a flexible to plastic, dextrinoid inner layer in the innermost germinal wall in most Acaulospora spp. (vs. no such structures have been found in spores of Ambispora spp.). Finally, only Am. gerdemannii is a dimorphic fungus, forming both acaulosporioid and glomoid spores. The glomoid morphotype of Am. gerdemannii has not so far been found in Poland and the only report of its existence is that of Morton and Redecker (2001).
REFERENCES
Allen M. F., MacMahon J. A. 1985. Impact of disturbance on cold desert fungi: comparative microscale dispersion patterns. Pedobiol. 28, 215-224.
An Z.-Q., Grove J. H., Hendrix J. W., Hershman D. E., Henson G. T. 1990. Vertical distribution of endogonaceous mycorrhizal fungi associated with soybean as affected by soil fumigation. Soil Biol. Biochem. 22, 715-719.
An Z.-Q., Guo B. Z., Hendrix J. W. 1993. Mycorrhizal pathogen of tobacco: cropping history and current crop effects on the mycorrhizal fungal community. Crop prot. 12, 527-531.
An Z.-Q., Hendrix J. W., Hershamn D. E., Ferriss R. S., Henson G. T. 1993. The influence of crop rotation and soil fumigation on a mycorrhizal fungal community associated with soybean. Mycorrhiza 3, 171-182.
An Z. Q., Quo B. Z., Hendrix J. W. 1993. Populations of spores and propagules of mycorrhizal fungi in relation to the life cycles of tall fescue and tobacco. Soil Biol. Biochem. 25, 813-817.
Bever J., Morton J. B., Antonovics J., Schultz P. A. 1996. Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. J. Ecol. 84, 71-82.
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone and Complexipes spec ies deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland.http://www.agro.ar.szczecin.pl/~jblaszkowski/.
Dodd J. C., Arias I., Koomen I., Hayman D. S. 1990. The management of populations of vesicualr-arbuscular mycorrhizal fungi in acid-infertile soils of a savanna ecosystem. Plant and Soil 122, 241-247.
Koske R. E., Gemma J. N., Jackson N. 1977. Mycorrhizal fungi associated with three species of turfgrass. Can. J. Bot. 75, 320-332.
Moreira-Souza M., Trufem F. B., Gomes-da-Costa S. M., Cardoso E. J. B. N. 2003. Arbuscular mycorrhizal fungi associated with Araucaria angustifolia (Bert.) O. Ktze. Mycorrhiza 13, 211-215.
Morton J. B. 2002. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University: http://www.invam.caf.wvu.edu/.
Morton J. B., Redecker D. 2001. Two families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus , based on concordant molecular and morphological characters. Mycologia 93, 181-195.
Nicolson T. H., Schenck N. C. 1979. Endogonaceous mycorrhizal endophytes in Florida. Mycologia 71, 178-198.
Rose S., Daniels B. A., Trappe J. M. 1979. Glomus gerdemannii sp. nov. Mycotaxon 8, 297-301.
Spain J. L., Sieverding E., Oehl F. 2006. Appendicispora: a new genus in the arbuscular mycorrhiza-forming Glomeromycetes, with a discussion of the genus Archaeospora. Mycotaxon 97, 163-182.
Turnau K., Ryszka P., Gianinazzi-Pearson V., Tuinen D. van 2001. Identification of arbuscular mycorrhizal fungi in soils and roots of plants colonizing zinc wastes in southern Poland. Mycorrhiza 10, 169-174.
Walker C. 1986. Taxonomic concepts in the Endogonaceae. II. A fifth morphological wall type in endogonaceous spores. Mycotaxon 25, 95-99.